

How Is Embryology Used As Evidence For Evolution
Charles Darwin's theory of evolution restructured comparative embryology and gave information technology a new focus. After reading Johannes Müller's summary of von Baer'south laws in 1842, Darwin saw that embryonic resemblances would exist a very stiff argument in favor of the genetic connectedness of different beast groups. "Community of embryonic structure reveals community of descent," he would conclude in On the Origin of Species in 1859.
Larval forms had been used for taxonomic classification even earlier Darwin. J. V. Thompson, for example, had demonstrated that larval barnacles were almost identical to larval venereal, and he therefore counted barnacles as arthropods, not molluscs (Effigy ane.12; Winsor 1969). Darwin, an expert on barnacle taxonomy, celebrated this finding: "Even the illustrious Cuvier did not perceive that a barnacle is a crustacean, but a glance at the larva shows this in an unmistakable manner." Darwin'south evolutionary estimation of von Baer'south laws established a paradigm that was to be followed for many decades, namely, that relationships betwixt groups can be discovered by finding common embryonic or larval forms. Kowalevsky (1871) would soon make a similar blazon of discovery (publicized in Darwin'due south Descent of Human being) that tunicate larvae have notochords and grade their neural tubes and other organs in a mode very like to that of the primitive chordate Amphioxus. The tunicates, some other enigma of nomenclature schemes (formerly placed, along with barnacles, among the molluscs), thereby found a home with the chordates.

Effigy 1.12
Nauplius larvae of (A) a barnacle (Tetraclita, seen in ventral view) and (B) a shrimp (Penaeus, seen in dorsal view). The shrimp and barnacle share a like larval phase despite their radical divergence in later development. (Afterwards Müller 1864.) (more than...)
Darwin as well noted that embryonic organisms sometimes make structures that are inappropriate for their adult class only that show their relatedness to other animals. He pointed out the being of eyes in embryonic moles, pelvic rudiments in embryonic snakes, and teeth in embryonic baleen whales.
Darwin as well argued that adaptations that depart from the "type" and allow an organism to survive in its detail environs develop late in the embryo.* He noted that the differences between species inside genera become greater equally development persists, every bit predicted by von Baer'southward laws. Thus, Darwin recognized 2 ways of looking at "descent with modification." One could emphasize the common descent past pointing out embryonic similarities betwixt two or more than groups of animals, or one could emphasize the modifications past showing how evolution was altered to produce structures that enabled animals to arrange to particular conditions.
Embryonic homologies
One of the most important distinctions made by the evolutionary embryologists was the departure between analogy and homology. Both terms refer to structures that appear to exist similar. Homologous structures are those organs whose underlying similarity arises from their existence derived from a mutual ancestral structure. For example, the wing of a bird and the forelimb of a man are homologous. Moreover, their respective parts are homologous (Figure 1.13). Analogous structures are those whose similarity comes from their performing a like role, rather than their arising from a common ancestor. Therefore, for instance, the wing of a butterfly and the wing of a bird are coordinating. The two types of wings share a common part (and therefore are both called wings), just the bird wing and insect wing did not ascend from an original ancestral structure that became modified through development into bird wings and butterfly wings.
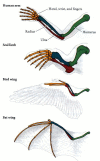
Effigy one.thirteen
Homologies of structure amongst a human being arm, a seal forelimb, a bird wing, and a bat wing; homologous supporting structures are shown in the same color. All four are homologous as forelimbs and were derived from a common tetrapod ancestor. The adaptations (more...)
Homologies must be fabricated carefully and must always refer to the level of organization being compared. For instance, the bird wing and the bat wing are homologous as forelimbs, but not every bit wings. In other words, they share a common underlying structure of forelimb bones because birds and mammals share a common beginnings. However, the bird wing developed independently from the bat wing. Bats descended from a long line of nonwinged mammals, and the structure of the bat wing is markedly different from that of a bird fly.
One of the most celebrated cases of embryonic homology is that of the fish gill cartilage, the reptilian jaw, and the mammalian middle ear (reviewed in Gould 1990). Commencement, the gill arches of jawless (agnathan) fishes became modified to class the jaw of the jawed fishes. In the jawless fishes, a series of gills opened behind the jawless oral cavity. When the gill slits became supported by cartilaginous elements, the outset set of these gill supports surrounded the mouth to class the jaw. There is ample evidence that jaws are modified gill supports. Commencement, both these sets of basic are fabricated from neural crest cells. (Most other bones come from mesodermal tissue.) Second, both structures form from upper and lower bars that bend forward and are hinged in the middle. Third, the jaw musculature seems to exist homologous to the original gill back up musculature. Thus, the vertebrate jaw appears to be homologous to the gill arches of jawless fishes.
Just the story does non end here. The upper portion of the second embryonic arch supporting the gill became the hyomandibular os of jawed fishes. This element supports the skull and links the jaw to the attic (Figure one.14A). Equally vertebrates came up onto land, they had a new problem: how to hear in a medium every bit thin as air. The hyomandibular bone happens to be near the otic (ear) capsule, and bony material is fantabulous for transmitting sound. Thus, while however functioning as a cranial caryatid, the hyomandibular bone of the commencement amphibians as well began functioning as a audio transducer (Clack 1989). Equally the terrestrial vertebrates contradistinct their locomotion, jaw structure, and posture, the attic became firmly attached to the rest of the skull and did not demand the hyomandibular caryatid. The hyomandibular bone and so seems to accept become specialized into the stapes bone of the middle ear. What had been this bone's secondary function became its master function.
The original jaw bones changed also. The first embryonic curvation generates the jaw apparatus. In amphibians, reptiles, and birds, the posterior portion of this cartilage forms the quadrate os of the upper jaw and the articular bone of the lower jaw. These bones connect to each other and are responsible for articulating the upper and lower jaws. Yet, in mammals, this articulation occurs at another region (the dentary and squamosal basic), thereby "freeing" these bony elements to learn new functions. The quadrate bone of the reptilian upper jaw evolved into the mammalian incus os of the center ear, and the articular bone of the reptile's lower jaw has become our malleus. This latter procedure was first described by Reichert in 1837, when he observed in the squealer embryo that the mandible (jawbone) ossifies on the side of Meckel's cartilage, while the posterior region of Meckel's cartilage ossifies, detaches from the residue of the cartilage, and enters the region of the centre ear to become the malleus (Figure 1.14B,C). Thus, the middle ear bones of the mammal are homologous to the posterior lower jaw of the reptile and to the gill arches of agnathan fishes. Chapter 22 will detail more recent information concerning the human relationship of development to evolution.
- *
-
Moreover, as first noted by Weismann (1875), larvae must have their own adaptations to help them survive. The adult viceroy butterfly mimics the monarch butterfly, but the viceroy caterpillar does not resemble the cute larva of the monarch. Rather, the viceroy larva escapes detection by resembling bird droppings (Begon et al. 1986).


How Is Embryology Used As Evidence For Evolution,
Source: https://www.ncbi.nlm.nih.gov/books/NBK10049/
Posted by: simonsdecten.blogspot.com
0 Response to "How Is Embryology Used As Evidence For Evolution"
Post a Comment